
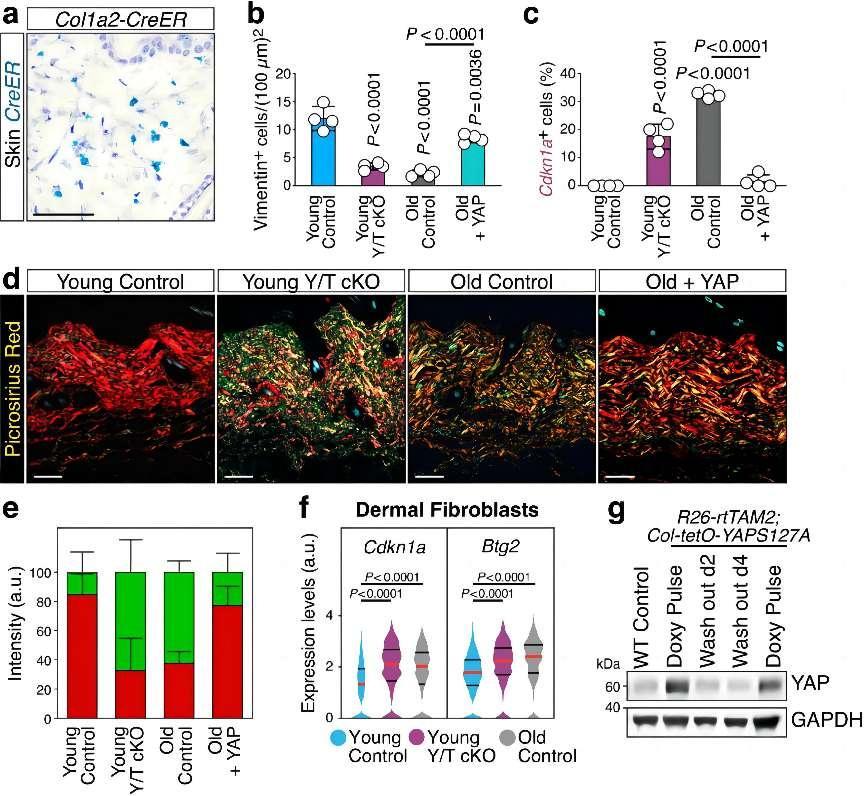
a, Representative in situ hybridization for Cre recombinase (CreER) in dorsal skin sections of a Col1a2-CreER mouse, showing specific staining in dermal fibroblasts (Scale bar, 50 µm). b, Quantification of Vimentin-positive cells in the dermal layer of skin sections from mice of the indicated genotypes. Data (n = 4 mice for each column) are presented as mean ± s.d.; P values are derived from one-way ANOVA with Dunnett’s multiple comparison test. Related to Fig. 2b. c, Quantification of Cdkn1a-positive cells in the dermal layer of skin sections from mice of the indicated genotypes. Data (n = 4 per condition) are presented as mean ± s.d.; P values are derived from one-way ANOVA with Dunnett’s multiple comparison test. Related to Fig. 2c. d, e, Representative birefringence images (d, Scale bar, 100 µm) and quantification (e) of picrosirius red staining on dorsal skin sections from mice of the indicated genotypes. In d, the red-to-green gradient reflects the decreasing thickness of collagen fiber bundles. In e, red is the quantification of thicker collagen fibers appearing as red or yellow in birefringence; green is the quantification of thinner fibers appearing as green in the same conditions. Data in e (n = 4 mice for each column) are presented as mean + s.d., 25 areas were analyzed per condition. f, Violin plots from sc-RNA seq data showing the expression distribution of Cdkn1a and Btg2 in freshly dissociated dermal fibroblasts of young control (n = 2916 cells), young Y/T cKO (n = 759 cells) and old control (n = 1413 cells) mice. Red lines denote the median, black lines denote quartiles. P values are derived from Kruskal-Wallis test. g, Immunoblotting analysis of YAP in tail tip biopsies from wild-type (WT Control) and R26-rtTAM2; Col-tetO-YAPS127A mice treated with intermittent doxycycline regime to sustain YAP activity. Mice were analyzed under doxycycline (Doxy Pulse) or during the off-doxy period, two or four days after the last doxy pulse (Wash out d2 and Wash out d4, respectively; see Methods). GAPDH served as loading control.
Credit:YAP/TAZ activity in stromal cells prevents ageing by controlling cGAS–STING.
YAP/TAZ are a class of transcription co-activators that play a key role in various cell types. Under normal physiological conditions, they regulate cell proliferation and apoptosis by modulating the Hippo signaling pathway, thereby maintaining tissue homeostasis. However, during tumorigenesis, the expression and activity of YAP/TAZ are abnormally upregulated, leading to uncontrolled cell proliferation and promoting tumor development. In addition to regulation of tissue homeostasis and tumorigenesis, a groundbreaking study published in Nature uncovered the pivotal role of YAP/TAZ in preventing premature aging and tissue dysfunction, offering a novel perspective on the molecular mechanisms underlying these processes. By using advanced molecular and cellular techniques, the researchers demonstrated that the decline in YAP/TAZ activity in stromal cells during natural aging correlated with the onset of senescence and age-related pathologies.
Firstly, the study revealed that YAP/TAZ activity diminished with age in stromal cells, such as fibroblasts and vascular smooth muscle cells. This decline was observed not only at the protein level but also in their subcellular localization. Furthermore, the study demonstrated that the genetic inactivation of YAP/TAZ in young mice accelerated aging, mimicking the natural aging process. These findings suggested that YAP/TAZ in stromal cells was crucial for maintaining tissue homeostasis and preventing premature aging.
Secondly, the study elucidated that sustaining YAP/TAZ activity could rescue the premature aging traits induced by YAP/TAZ inactivation. For instance, YAP/TAZ knockout mice exhibited features of aging, such as reduced fibroblast density, upregulation of senescence-associated genes, and abnormal collagen deposition. However, these aging traits could be reversed by reintroducing active YAP/TAZ. This demonstrated that YAP/TAZ in stromal cells played a key role in preventing premature aging.
Thirdly, the study unveiled that YAP/TAZ activity was essential for maintaining nuclear envelope integrity. The inactivation of YAP/TAZ caused nuclear envelope defects and nuclear herniations, which were linked to the activation of cGAS-STING signaling. This signaling pathway has been shown to induce senescence and aging. Furthermore, the study identified that YAP/TAZ inhibited cGAS-STING signaling by regulating the expression of lamin B1 and ACTR2, which were required for nuclear envelope integrity and the formation of the perinuclear actin cap. This role of YAP/TAZ in maintaining nuclear envelope integrity was crucial for suppressing cGAS-STING signaling. Subsequently, the study showed that inhibiting cGAS-STING signaling in YAP/TAZ cKO mice prevented the accumulation of senescent cells and premature aging traits. This indicated that the activation of cGAS-STING signaling downstream of YAP/TAZ inactivation drove senescence and aging.
In conclusion, the study provided new insights into the molecular mechanisms by which YAP/TAZ maintained tissue integrity and prevented premature aging, with implications for the development of aging-related diseases. The findings suggested that YAP/TAZ functioned to preserve tissue homeostasis and prevent aging by controlling cGAS-STING signaling. Inhibition of STING or activation of YAP/TAZ might be effective in limiting senescence-associated inflammation and promoting healthy aging.
Although YAP/TAZ is important for preventing age-related diseases, its activation may be associated with the development of organ fibrosis in some cases. Fibrosis is a common feature of chronic diseases that involves the excessive production of extracellular matrix proteins, leading to scar tissue formation. Transforming growth factor-β (TGF-β) is known to be a key driver of fibrosis, and the transcription factors YAP and TAZ are also involved. However, the mechanism linking tissue injury to the activation of these transcription factors has not been fully understood.
Another study, conducted by Zhang et al., explored the role of the gene NUAK1 in promoting organ fibrosis and its crosstalk with TGF-β and YAP/TAZ. NUAK1 belongs to the AMPK family and is a class of serine/threonine protein kinases. The researchers identified that the gene NUAK1 was upregulated in fibrotic human kidney tissue and in mouse models of kidney, liver, and lung fibrosis. They found that NUAK1 expression was regulated by TGF-β, YAP, and TAZ. Mice lacking the gene Nuak1 in fibroblasts were protected from fibrosis, and treatment with a NUAK1 inhibitor also reduced fibrosis. The authors demonstrated that NUAK1 was induced by TGF-β and promoted YAP and TGF-β/SMAD signaling to drive fibrogenesis, indicating NUAK1 as a potential therapeutic target.
The study first profiled the transcriptomes of human kidney biopsy samples with varying degrees of fibrosis, revealing NUAK1 as a gene highly correlated with fibrosis. Subsequent analysis of mouse models of fibrosis confirmed the upregulation of Nuak1 in fibrotic organs. Mechanistic experiments showed that TGF-β induced NUAK1 expression in fibroblasts, which in turn promoted YAP activation and expression of fibrosis-related genes.
Importantly, the study identified a positive feedback loop between NUAK1 and YAP/TAZ. NUAK1 promoted YAP and TGF-β/SMAD signaling, which induced further NUAK1 expression. This loop perpetuated fibroblast activation and fibrosis progression. Furthermore, fibroblast-specific deletion of Nuak1 in mice significantly reduced fibrosis in kidney, lung, and liver fibrosis models, indicating the critical role of fibroblast NUAK1 in fibrogenesis.
The study also demonstrated the potential of NUAK1 inhibition as an antifibrotic strategy. Treatment with a NUAK1 inhibitor starting immediately after injury reduced fibrosis development, and delayed initiation of treatment also attenuated established fibrosis. This suggested that NUAK1 inhibition could be beneficial even after fibrosis has developed.
In summary, this study identified NUAK1 as a key driver of fibrosis through its regulation of YAP and TGF-β/SMAD signaling. The NUAK1-YAP positive feedback loop sustained fibroblast activation and fibrosis progression. These findings supported the potential of NUAK1 inhibition as a novel antifibrotic therapy.
Sladitschek-Martens HL, Guarnieri A, Brumana G, Zanconato F, Battilana G, Xiccato RL, Panciera T, Forcato M, Bicciato S, Guzzardo V, Fassan M, Ulliana L, Gandin A, Tripodo C, Foiani M, Brusatin G, Cordenonsi M, Piccolo S. YAP/TAZ activity in stromal cells prevents ageing by controlling cGAS-STING. Nature. 2022 Jul;607(7920):790-798.
Zhang T, He X, Caldwell L, Goru SK, Ulloa Severino L, Tolosa MF, Misra PS, McEvoy CM, Christova T, Liu Y, Atin C, Zhang J, Hu C, Vukosa N, Chen X, Krizova A, Kirpalani A, Gregorieff A, Ni R, Chan K, Gill MK, Attisano L, Wrana JL, Yuen DA. NUAK1 promotes organ fibrosis via YAP and TGF-β/SMAD signaling. Sci Transl Med. 2022 Mar 23;14(637):eaaz4028.






Post comments