
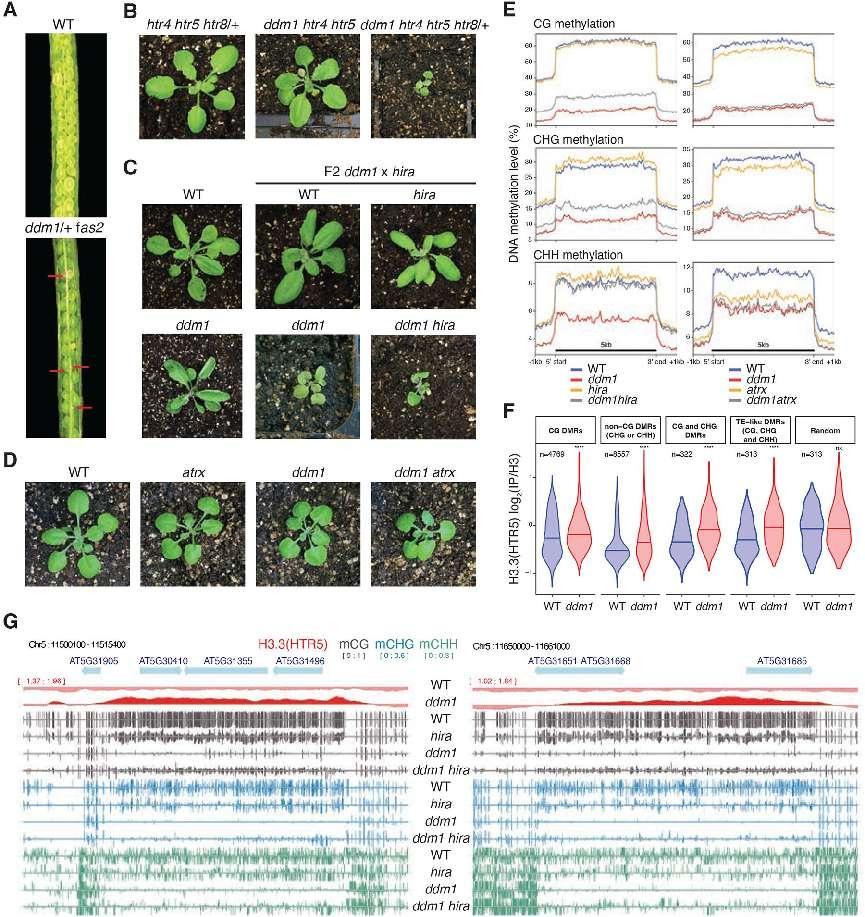
Genetic interactions between ddm1 and histone H3 variants and chaperones impact DNA methylation.
fas2 and hira are mutants in H3.1 (CAF-1), and H3.3 (HIRA) chaperones, respectively. ATRX is a chromatin remodeler required for H3.3 deposition. (A) Siliques of wild-type (WT) and ddm1/+ fas2 plants. Red arrows indicate nonviable seeds (synthetic lethality). (B) F2 ddm1 htr4 htr5 htr8/+ with reduced H3.3 has severe growth phenotypes compared to htr4 htr5 htr8/+. (C) F2 ddm1 hira double mutants from ddm1 and hira parents, compared with WT, hira and ddm1 siblings. (D) F2 ddm1 atrx double mutants from ddm1 and atrx parents compared with WT and ddm1 siblings. ddm1 hira and ddm1 atrx were phenotypically indistinguishable from ddm1 siblings but ddm1 hira were more severe. (E) DNA methylation levels in CG, CHG and CHH contexts in ddm1 and hira mutants on the left, and ddm1 and atrx mutants on the right, determined by whole genome bisulfite sequencing. The DNA methylation levels range from 0 to 100% and are substantially increased in ddm1 hira as compared to ddm1. atrx mutants lose some methylation and fail to rescue methylation loss in ddm1. Metaplots calculated from all 31,189 transposable elements annotated in TAIR10. (F) Levels of H3.3 in WT and ddm1 Chip-seq at differentially methylated regions (DMRs) between ddm1 and ddm1 hira (hyper-methylated in ddm1 hira). The number of DMRs (n) in the different cytosine nucleotide contexts are noted. H3.3 is statistically enriched in ddm1 compared to WT at these DMRs, but not in random regions (**** P<0.0001, ns not significant, t-test). See Table S1 for the list of all DMRs. (G) Representative loci that re-gain DNA methylation in ddm1 hira as compared to ddm1. Ectopic H3.3 in ddm1 is shown above (red track).
Credit:DOI:10.1016/j.cell.2023.08.001
A recent article published in Cell delved into the molecular mechanism of chromatin remodeling by DECREASE in DNA METHYLATION 1 (DDM1) underlying epigenetic inheritance of DNA methylation. DDM1 is a chromatin remodeling factor that plays a crucial role in maintaining the epigenetic state of DNA methylation. It is involved in remodeling chromatin during the cell cycle to facilitate access of DNA methyltransferases to heterochromatin. DDM1 is essential for the epigenetic inheritance of DNA methylation, as it allows DNA methylation of transposons in the male germline. Clinically, DDM1 has been associated with several diseases. It is involved in immunodeficiency-centromeric instability-facial anomalies (ICF) syndrome, a rare autosomal recessive disorder characterized by immune deficiency and centromere instability. Mutations in DDM1 have also been linked to autoimmune diseases and cancer.
The authors highlighted the crucial role of the chromatin remodeler DDM1 in preferentially remodeling heterochromatin nucleosomes during S phase, allowing DNA methylation by the methyltransferase MET1. This process involved the replacement of histone variant H3.3 by H3.1, facilitated by DDM1, with subsequent deposition of H2A.W stabilizing the H3.1 nucleosomes and permitting DNA methylation.
The study's key findings included the discovery of an N-terminal autoinhibitory domain in DDM1 that controlled its remodeling activity, enabling preferential targeting of H3.3 nucleosomes. The structure of DDM1 bound to a variant nucleosome reveals direct contacts with H3.3, indicating a mechanism for specific histone variant recognition. Furthermore, the male germline H3.3 variant MGH3 acted as a placeholder nucleosome, resisting DDM1 remodeling and facilitating transgenerational inheritance of unmethylated transposons.
This research provided a molecular model for epigenetic inheritance of DNA methylation where DDM1 played a crucial role in maintaining the epigenetic state of DNA methylation and had significant implications in both normal biology and disease. Its involvement in chromatin remodeling and epigenetic inheritance made it an important target for research and potential therapeutic interventions. For example, it has been shown to promote tumor growth and metastasis in various types of cancer. Inhibition of DDM1 may have therapeutic benefits by disrupting the epigenetic regulation of gene expression in cancer cells.
In addition to DNA methylation, which is a core regulatory mechanism in epigenetics, chemical modifications of histones, such as methylation and acetylation, are also important research topics in this field. These modifications act on histones, the proteins around which DNA is wrapped in nucleosomes, and alter their chemical properties, which in turn affect the packaging and readability of DNA. This allows for the regulation of gene expression without altering the gene DNA sequence itself. DNA methylation is typically associated with gene silencing and the condensation of chromatin structure, while histone acetylation is related to gene activation and the opening of chromatin structure. These complex epigenetic regulatory mechanisms play key roles in biological processes such as cell differentiation, development, the occurrence of diseases, and responses to environmental changes.
Another article published in Nature identified a novel post-translational modification, Nε-acetyl-Nε-methyllysine (Kacme), found on histone H4 in multiple species and tissues. This modification was enriched at the promoters of actively transcribed genes and was dynamically regulated in response to biological signals. The discovery demonstrated that histone lysine methylation and acetylation could coexist at the same residue, challenging the previous assumption of their mutual exclusivity. The study identified the acetyltransferase p300 as a writer of Kacme, which was installed through the sequential modification of monomethylated lysine residues. Additionally, it showed that Kacme was recognized by bromodomain proteins like BRD2, enabling its regulatory role in chromatin. The results established Kacme as a distinct regulatory modification beyond acetylation and methylation alone, with potential roles in chromatin biology and gene expression regulation. This finding expanded the understanding of histone modifications and their impacts on chromatin dynamics and transcription, presenting new opportunities for studying epigenetic regulation and gene expression control.
Based on the complicated crosstalk between histone methylation and acetylation, Jain et al. investigated the regulatory mechanism of H3K4 methylation by acetylation of the H3 tail in nucleosomes. They found H3 tail acetylation enhanced the binding of PHD finger reader domains to nucleosomal H3K4me3, and increased the methylation activity of the methyltransferase MLL1 core complex (MLL1C) towards nucleosomal H3K4. Besides, H3K4me3 preferentially co-occurred on highly acetylated H3 tails, as observed in vivo using mass spectrometry of mammalian cells. This correlation strengthened with acetylation inhibitor treatment.
The study proposed a model where H3 tail acetylation promoted nucleosome accessibility, facilitating H3K4me3 engagement by readers and writers. This resolved the long-standing question of why H3K4me3 and H3 acetylation were coupled. The study's findings indicated a critical role of H3 tail acetylation in regulating H3K4 methylation and epigenetic gene regulation. This acetylation-mediated chromatin switch was suggested to control the read and write access of the H3 tail, thereby governing H3K4 methylation levels in cells. Overall, the study provided valuable insights into the cross-talk between histone modifications and their regulatory roles in epigenetic processes.
Lee SC, Adams DW, Ipsaro JJ, Cahn J, Lynn J, Kim HS, Berube B, Major V, Calarco JP, LeBlanc C, Bhattacharjee S, Ramu U, Grimanelli D, Jacob Y, Voigt P, Joshua-Tor L, Martienssen RA. Chromatin remodeling of histone H3 variants by DDM1 underlies epigenetic inheritance of DNA methylation. Cell. 2023 Sep 14;186(19):4100-4116.e15.
Lu-Culligan WJ, Connor LJ, Xie Y, Ekundayo BE, Rose BT, Machyna M, Pintado-Urbanc AP, Zimmer JT, Vock IW, Bhanu NV, King MC, Garcia BA, Bleichert F, Simon MD. Acetyl-methyllysine marks chromatin at active transcription start sites. Nature. 2023 Oct;622(7981):173-179.
Jain K, Marunde MR, Burg JM, Gloor SL, Joseph FM, Poncha KF, Gillespie ZB, Rodriguez KL, Popova IK, Hall NW, Vaidya A, Howard SA, Taylor HF, Mukhsinova L, Onuoha UC, Patteson EF, Cooke SW, Taylor BC, Weinzapfel EN, Cheek MA, Meiners MJ, Fox GC, Namitz KEW, Cowles MW, Krajewski K, Sun ZW, Cosgrove MS, Young NL, Keogh MC, Strahl BD. An acetylation-mediated chromatin switch governs H3K4 methylation read-write capability. Elife. 2023 May 19;12:e82596.






Post comments