
Credit:Head-to-head comparison of relevant cell sources of small extracellular vesicles for cardiac repair: Superiority of embryonic stem cells
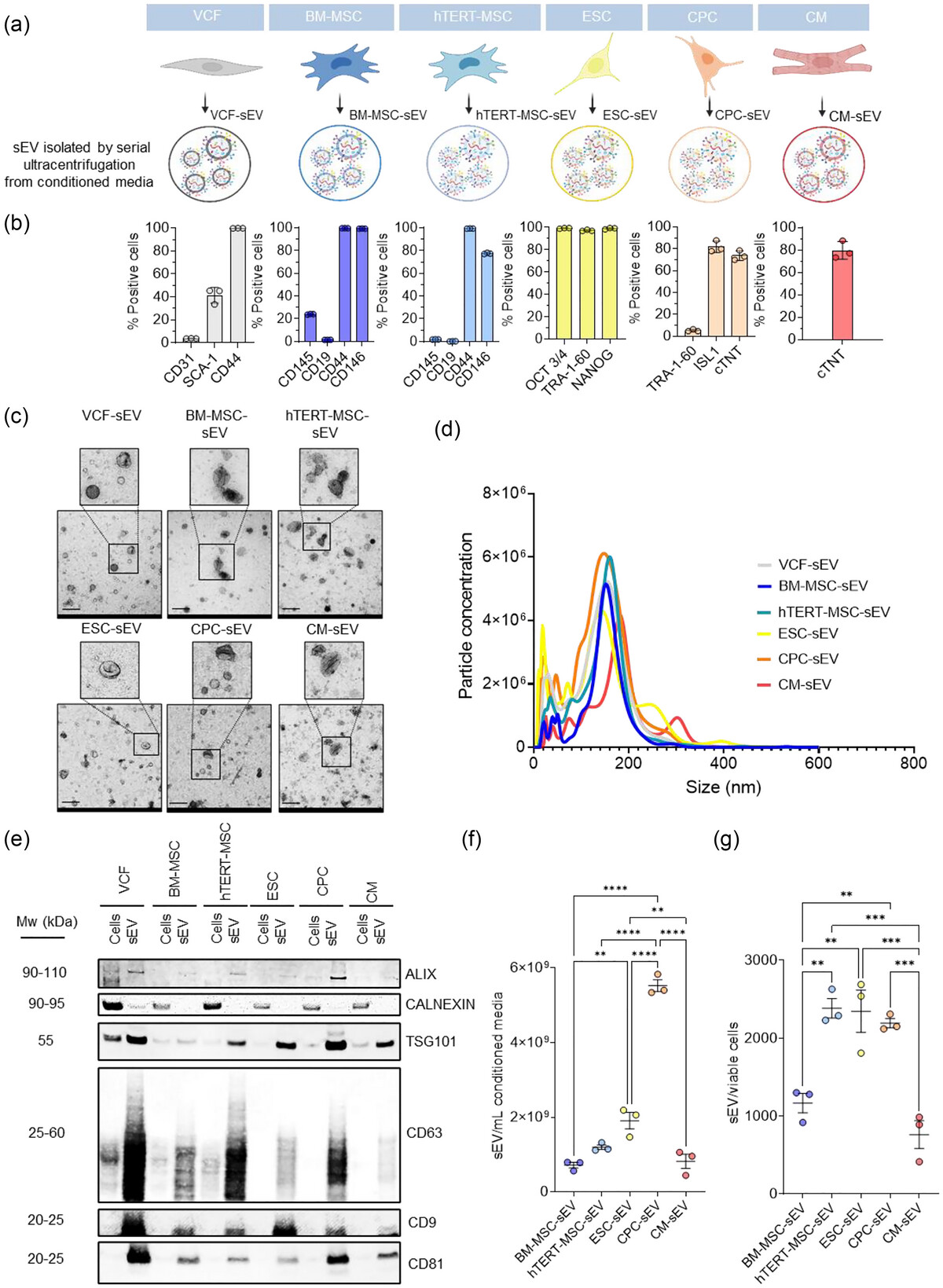
Characterizations of vesicles isolated from various cell sources. (a) Schematic representation of the different cell sources used for sEV preparation. (b) Bar graphs showing the percentage of cells expressing each indicated cell type-specific marker measured by flow cytometry. (c) Electron microscopy images of isolated sEV. Enlarged images highlight detailed vesicles morphology. Scale bar: 200 nm. (d) sEV size distribution analysis with the NanoSight LM14c. (e) Western blot analysis for ALIX, CALNEXIN, TSG101, CD63, CD9 and CD81 in protein extracts from indicated sEV and their parental cells. Five µg of total cell or sEV protein was loaded in the Western blot for every sample. (f) sEV yield per mL of conditioned media from indicated cell sources. (g) sEV yield per viable cell from the indicated cell sources. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. One-way ANOVA followed by Tukey post-hoc test were applied. Dots in the bar graphs represent biological replicates. N = 3. MW (Molecular weight).
Following myocardial infarction (MI), a substantial number of cardiomyocytes undergo apoptosis, leading to the formation of fibrotic scars that impede cardiac repair and functional recovery. While stem cell therapy has demonstrated some promise in cardiac regeneration, its clinical application is fraught with challenges such as low survival rates of transplanted cells, immune rejection, and potential tumorigenicity. Moreover, the migration and engraftment of stem cells within the host tissue are challenging to control, further diminishing their therapeutic efficacy. Recently, small extracellular vesicles (sEVs) have emerged as significant players in cardiac repair. Nevertheless, sEVs derived from different sources may vary considerably in their functional properties and effectiveness.
González-King et al. systematically compared the effects of small extracellular vesicles (sEVs) derived from various sources, including human bone marrow mesenchymal stem cells (BM-MSCs), human immortalized MSCs (hTERT-MSCs), human embryonic stem cells (ESCs), ESC-derived cardiac progenitor cells (CPCs), cardiomyocytes (CMs), and human ventricular cardiac fibroblasts (VCFs), in cardiac repair. They identified the most promising source of sEVs and elucidated their specific mechanisms of action. sEVs were initially extracted from these cells using ultracentrifugation and subsequently evaluated through a series of in vitro and in vivo models for their roles in angiogenesis, fibrosis, cardioprotection, cell proliferation, and macrophage response regulation.
In the in vitro cardiac repair function experiments, sEVs derived from hTERT-MSCs, BM-MSCs, CPCs, and CMs significantly reduced cardiomyocyte death by approximately two-fold, whereas ESC-derived sEVs did not show a significant effect. Furthermore, ESC-derived sEVs exhibited the strongest pro-angiogenic ability in both capillary-like tube formation and aortic ring sprouting assays and significantly reduced the expression of mature collagen and α-SMA in VCFs. These results indicate that ESC-derived sEVs are most effective in promoting angiogenesis and anti-fibrosis, while sEVs from other sources also demonstrate efficacy in cardioprotection. Regarding macrophage response regulation, the experiments revealed that sEVs derived from BM-MSCs, ESCs, and CPCs promoted the transition of macrophages from the M1 to M2 phenotype and reduced the secretion of inflammatory cytokines IL-6 and TNF-α by M1 macrophages.
Importantly, ESC-derived sEVs also demonstrated significant cardiac repair effects in a mouse model of myocardial infarction-reperfusion injury, particularly by reducing fibrosis and promoting angiogenesis. Multi-omics analysis revealed that genes and proteins specifically upregulated in ESC-derived sEVs were primarily involved in angiogenesis and cell growth pathways, such as ROBO, BDNF, MYC, and telomerase-related pathways, while fibrosis-related pathways, including collagen metabolism, WNT, TGF-β, and PDGF signaling pathways, were downregulated. These findings corroborate the results from both in vitro and in vivo experiments, elucidating the molecular mechanisms underlying the superior cardiac repair capabilities of ESC-derived sEVs.
In conclusion, the study found that ESC-derived sEVs are most effective in promoting angiogenesis and anti-fibrosis, demonstrating significant potential for cardiac repair. ESC-derived sEVs can exert substantial cardiac repair effects by directly regulating angiogenesis and fibrosis and indirectly modulating macrophage phenotypes. In the future, ESC-derived sEVs could serve as a promising acellular therapeutic approach for cardiac repair post-myocardial infarction, warranting further research and development.
However, despite the significant contributions of this study in comparing the effectiveness of sEVs from different cell sources in cardiac repair, several limitations and questions remain that require further investigation. These include understanding the selective molecular transfer mechanisms of sEVs, determining optimal dosages and delivery routes, and evaluating the effects of sEVs under various disease conditions. Future research should focus on deeper mechanistic analyses, standardization of production processes, optimization of sEV sources, clinical translation, ethical and regulatory considerations, as well as the long-term efficacy and safety of sEVs. Such comprehensive exploration will advance sEV technology towards clinical applications.
González‐King, Hernán, et al. "Head‐to‐head comparison of relevant cell sources of small extracellular vesicles for cardiac repair: Superiority of embryonic stem cells." Journal of Extracellular Vesicles 13.5 (2024): e12445.






Post comments