
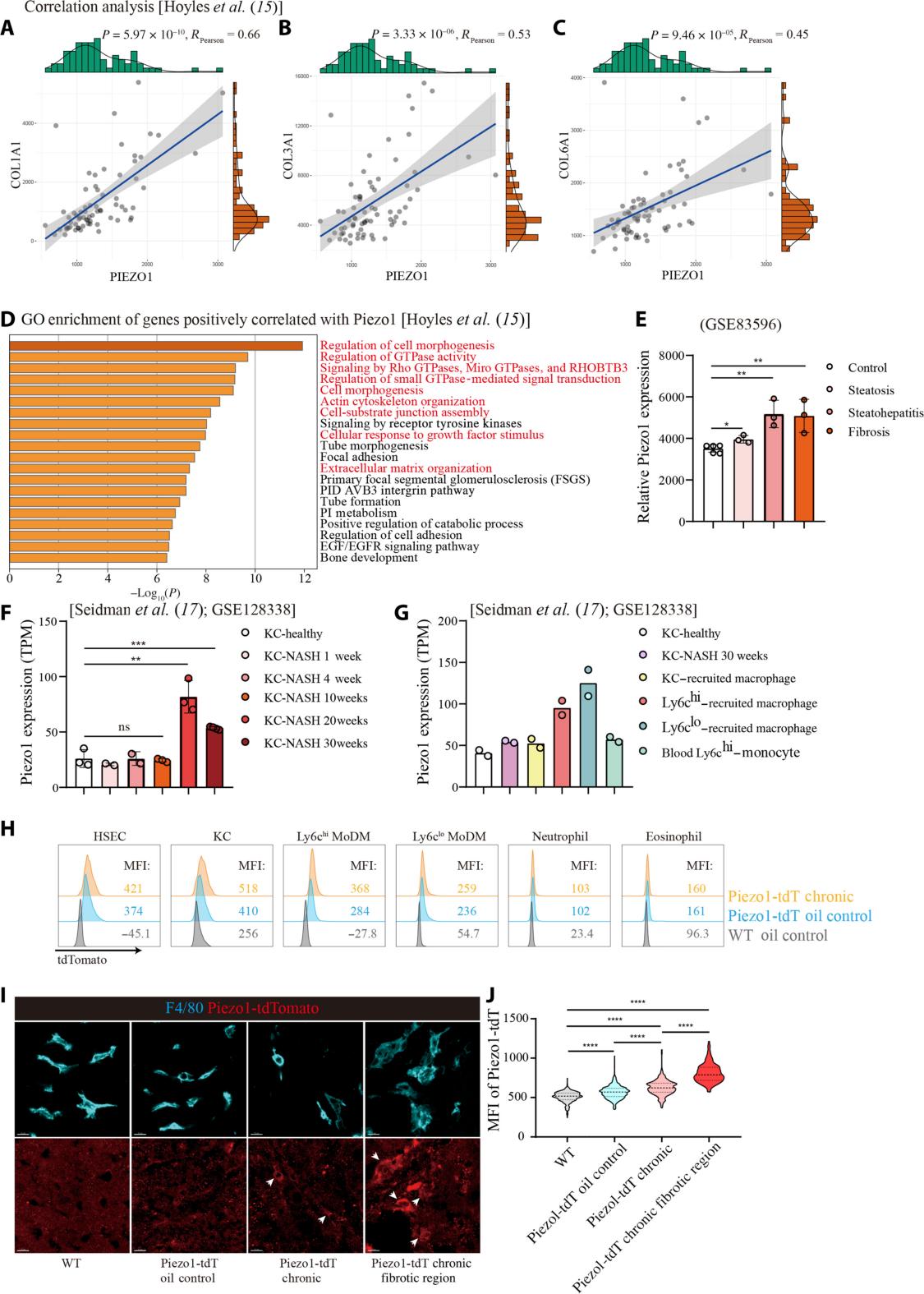
Increased expression of Piezo1 in liver macrophages during the progression of NASH.
(A to C) Correlation analysis of PIEZO1 with COL1A1 (A), COL3A1 (B), and COL6A1 (C) using human transcription profiling dataset from public data. Every spot represents an individual patient. The P values and correlation coefficients were calculated using Pearson’s correlation analysis. (D) Biofunction analysis of top 100 genes positively correlated with PIEZO1 in human transcription profiling dataset. Pathways mentioned in the text are highlighted. (E) Normalized Piezo1 expression in mice liver during NASH progression analyzed by transcription profiling dataset from public data. Each spot represents a biological duplication of samples. (F and G) Normalized Piezo1 expression in Kupffer cells (F) and different monocyte or macrophage subpopulations (G) during NASH progression analyzed by transcription profiling dataset from public datasets. Each spot represents a biological duplication of samples (E to G). (H) Histogram plots of tdTomato intensities of HSECs, KCs, Ly6chi MoDMs, Ly6clo MoDMs, neutrophils, and eosinophils measured by flow cytometry. The mean fluorescence intensity (MFI) of cells from wild-type mice liver (WT oil control), Piezo1P1tdT mice liver treated with corn oil (Piezo1-tdT oil control) or repetitive CCl4 challenge (Piezo1-tdT chronic) was shown in the corresponding colors. (I) Representative confocal images of liver sections showing tdTomato (red) expression in F4/80-positive (cyan) liver macrophages in WT mice, Piezo1-tdT oil control mice, normal region, and fibrotic region of Piezo1-tdT oil control mice. Scale bars, 10 μm. (J) Quantification of tdTomato expression in liver macrophages in (I). Data are displayed as means ± SD. Unpaired Student’s two-tailed t test (E and F). One-way analysis of variance (ANOVA) with Tukey’s multiple comparisons test (J). ns, nonsignificant, *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001. Data are representative of two independent experiments (I and J).
Liver fibrosis represents a pathological alteration in the liver's injury repair process, typically resulting from chronic liver disease. It is a leading cause of cirrhosis, liver failure, and hepatocellular carcinoma, posing a significant threat to patients' health and survival. Macrophages are pivotal immune cells involved in liver fibrosis, exhibiting dual functions. During the early stages of fibrosis, macrophages exacerbate inflammatory responses and fibrosis progression by secreting pro-inflammatory cytokines and chemokines. Conversely, in the later stages, macrophages may adopt an anti-inflammatory phenotype, thereby facilitating the resolution of inflammation and regression of fibrosis. Emerging evidence indicates that tissue mechanical properties, such as stiffness, substantially influence cell function and tissue repair. Piezo1, a mechanosensitive ion channel protein responsive to mechanical stimuli such as stretch and pressure, has been implicated in various physiological and pathological processes. However, its specific role in liver fibrosis remains poorly understood. Wang et al. investigate the role of Piezo1 in liver fibrosis, focusing on its regulation of macrophage phagocytic function and anti-inflammatory responses in response to tissue stiffness.
Initially, researchers established a mouse model of liver fibrosis through repeated intraperitoneal injections of carbon tetrachloride (CCl4). They then generated macrophage-specific Piezo1-deficient mice (Piezo1fl/fl Lyz2Cre)and compared them with control mice (Piezo1fl/fl). The study found that Piezo1 expression was significantly upregulated in the fibrotic livers of both humans and mice. Flow cytometry and immunofluorescence analyses revealed that Piezo1 expression was markedly increased in liver macrophages, hepatic sinusoidal endothelial cells, and neutrophils in fibrotic mice, with the highest levels observed in Ly6clo monocyte-derived macrophages (MoDMs).
In Piezo1fl/fl Lyz2Cre mice, the extent of liver fibrosis was significantly greater 48 hours after the final CCl4 injection, accompanied by elevated mRNA levels of fibrosis-related genes (α-SMA, Timp1, Col1a1, Col3a1, Col6a1). Serum levels of ALT and AST were also significantly higher in Piezo1-deficient mice, indicating exacerbated liver injury. During CCl4-induced chronic liver injury, Piezo1-deficient macrophages demonstrated a significantly reduced capacity to phagocytose apoptotic cells, particularly neutrophils.
In vitro experiments confirmed that Piezo1-deficient bone marrow-derived macrophages (BMDMs) exhibited decreased phagocytosis of apoptotic cells. When cultured on substrates of varying stiffness, macrophages showed enhanced phagocytic capacity with increasing substrate stiffness, whereas Piezo1-deficient macrophages did not exhibit this increased phagocytic ability on rigid substrates. Furthermore, treatment with the Piezo1-specific agonist Yoda1 significantly enhanced the phagocytic capacity of wild-type BMDMs but was ineffective in Piezo1-deficient BMDMs.
Notably, Piezo1-deficient macrophages showed impaired phagosome acidification, highlighting Piezo1's crucial role in the acidification process and digestion of apoptotic cargo following phagocytosis. RNA sequencing analysis revealed that Piezo1-deficient macrophages had upregulated expression of pro-inflammatory and ECM-related genes, while anti-inflammatory gene expression was downregulated.
In summary, this study confirms that Piezo1 plays a protective role in the resolution of liver fibrosis by sensing tissue stiffness. This process enhances macrophage phagocytosis and the clearance of apoptotic cells, thereby limiting pro-inflammatory responses. Additionally, systemic application of the Piezo1 agonist Yoda1 significantly alleviated liver fibrosis, suggesting that Piezo1 could serve as a potential therapeutic target for treating liver fibrosis.
Previous studies have primarily focused on chemical signals and molecular mechanisms, often neglecting the role of mechanical signals in the fibrosis process. This study highlights the critical role of the mechanosensitive ion channel Piezo1 in liver fibrosis, particularly its regulation of macrophage phagocytic function through the sensing of tissue stiffness. This insight could open up additional therapeutic targets and strategies. Unlike traditional flat culture methods, the substrate culture system employed in this study more accurately mimics the in vivo microenvironment, providing a better reflection of macrophage behavior during fibrosis. Additionally, this method facilitates the screening and evaluation of potential drugs. Notably, the use of Yoda1, as compared to traditional anti-fibrotic drugs, offers advantages in specificity and a well-defined mechanism of action.
Regrettably, this study has several limitations and aspects requiring further validation. For example, it does not elucidate the downstream signaling pathways activated by Piezo1, particularly how these pathways regulate macrophage phagocytic function and anti-inflammatory responses. Additionally, while the study underscores the importance of Piezo1 in sensing tissue stiffness for macrophage phagocytosis and anti-inflammatory responses, the specific mechanisms by which mechanical and chemical signals interact remain unclear.
In summary, future research should further investigate Piezo1’s functions in various diseases and cell types, as well as its potential and safety in clinical applications. Such studies will contribute to a more comprehensive understanding of Piezo1's role in fibrosis and immune responses, thereby providing a stronger foundation for the development of novel therapeutic strategies.
Wang, Yang, et al. "Stiffness sensing via Piezo1 enhances macrophage efferocytosis and promotes the resolution of liver fibrosis." Science Advances 10.23 (2024): eadj3289.





Post comments