

Credit:www.freepik.com
Metabolic reprogramming plays a crucial role in shaping cellular responses to various stimuli, including inflammation and infection. Recent studies have highlighted the complex interplay between metabolism and innate immunity, revealing unexpected roles for metabolites and metabolic enzymes in modulating immune responses. This review will explore the emerging evidence linking metabolic enzymes, particularly fumarate hydratase (FH), to innate immune activation and its implications for diseases such as hereditary leiomyomatosis and renal cell carcinoma (HLRCC) and systemic lupus erythematosus (SLE).
Fumarate, a metabolic intermediate of the tricarboxylic acid (TCA) cycle, has been traditionally associated with tumor progression and is considered an oncometabolite. However, accumulating evidence suggested that fumarate also played a crucial role in innate immunity. Two recent studies demonstrated that increased levels of intracellular fumarate could activate the innate immune response in both mice and humans.
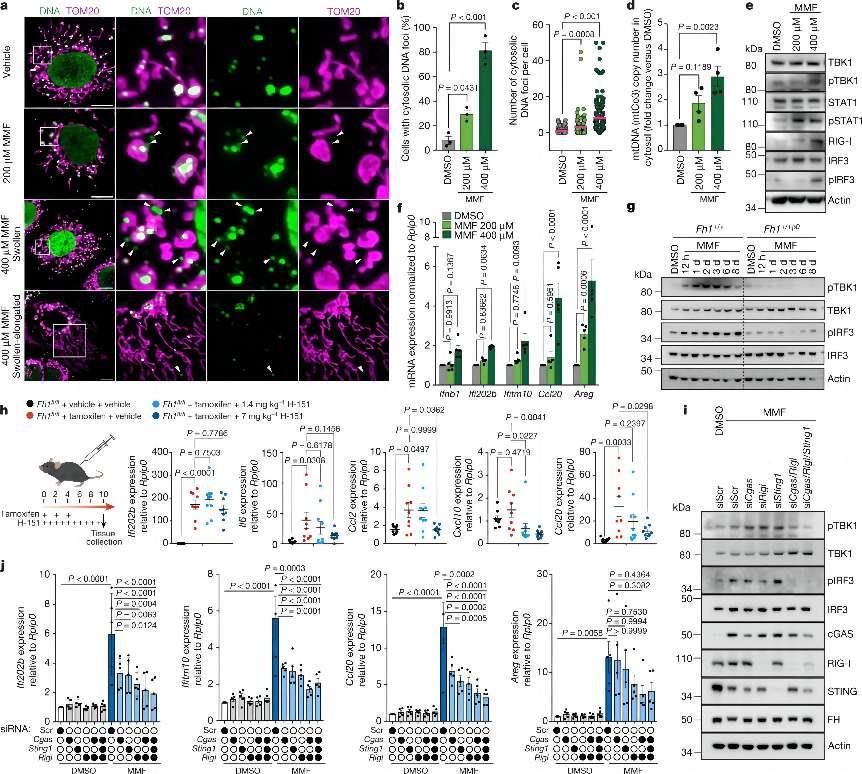
Fumarate induces a remodelling of mitochondrial morphology and the release of mtDNA.
a–f, Chronic Fh1 kidney cells (cFh1fl/fl) were treated with 200 μM or 400 μM MMF or vehicle (dimethyl sulfoxide; DMSO) for 8 days. n = 3 independent experiments (unless otherwise specified). In g, iFh1fl/flCL29 cells were treated with 400 μM MMF or DMSO for 8 days. a, Representative confocal images of mitochondrial morphology (TOM20) and DNA foci (DNA) in cFh1fl/fl cells. White arrowheads indicate cytosolic DNA foci. Scale bars, 10 μm. b,c, Percentage of cFh1fl/fl cells showing cytosolic DNA foci (b) and number of cytosolic DNA foci per cell (c) from a. d, Quantification of mtDNA copy number by ddPCR using a mtCo3 probe, from isolated cytosolic fractions of cFh1fl/fl cells. e, Immunoblots of specified proteins in cFh1fl/fl cells. f, ISG expression in cFh1fl/fl cells measured by qRT–PCR. n = 5 independent experiments. g, Immunoblots of specified proteins in iFh1fl/flCL29 and mtDNA-depleted iFh1fl/flCL29ρ0 cells. h, mRNA expression of a panel of ISGs in mouse kidney tissue treated with the STING inhibitor H-151, measured by qRT–PCR. n = 9 mice per group. i,j, Immunoblots of specified proteins (i) and ISG expression measured by qRT–PCR (j) in cFh1fl/fl cells treated with 400 µM MMF or DMSO for 8 d, and transfected with the indicated siRNAs (Scr, scramble). n = 5 independent experiments. Data are mean ± s.e.m. b–d,f,j, One-way ANOVA with Tukey’s multiple comparison test; h, one-way ANOVA with Dunnett’s multiple comparison test.
Credit:Fumarate induces vesicular release of mtDNA to drive innate immunity.
Zecchini et al. demonstrated that FH deficiency triggers early alterations in mitochondrial morphology, leading to the formation of swollen and swollen-elongated mitochondria. This was accompanied by the release of mitochondrial DNA (mtDNA) into the cytosol. The authors proposed that the accumulation of fumarate directly affected mitochondrial function, disrupting the balance between fusion and fission processes and ultimately causing mtDNA leakage.
The released cytosolic mtDNA acted as a sensor for the innate immune system. It triggered the activation of the cGAS-STING pathway, leading to the phosphorylation of IRF3 and STAT1 and the upregulation of interferon-stimulated genes (ISGs), which were key players in the immune response. Additionally, the study revealed a novel role for the RNA sensor RIG-I in mtDNA-induced innate immune activation. This dual sensing mechanism suggested a complex network of pathways mediating the immune response to mtDNA release.
The authors identified SNX9, a sorting nexin protein, as a critical mediator of mtDNA release. They demonstrated that SNX9 knockdown impaired mtDNA release and reduced the activation of the cGAS-STING pathway, indicating that SNX9-dependent mitochondrial-derived vesicles (MDVs) were responsible for delivering mtDNA to the cytosol. Further investigation revealed that these MDVs were selectively enriched in matrix proteins like pyruvate dehydrogenase (PDH) and contain TFAM, confirming their mitochondrial origin.
The study extended its findings to human HLRCC by analyzing gene expression profiles and inflammatory markers in FH-deficient renal cell carcinoma tissues. The results showed a striking similarity to the mouse model, with increased ISG expression, leukocyte infiltration, and elevated levels of pro-inflammatory cytokines like interleukin-6 (IL-6). This suggested that the fumarate-driven innate immune response is a key feature of human HLRCC.
Another interesting work conducted by Hooftman et al. investigated the role of FH in macrophage metabolism and immune responses. Using unbiased metabolomics and stable isotope tracing, the researchers uncovered an intriguing phenomenon: following stimulation with lipopolysaccharide (LPS), a component of bacterial cell walls, macrophages underwent metabolic reprogramming that led to the accumulation of fumarate. This accumulation was driven by increased expression of argininosuccinate synthase (ASS1), an enzyme that supported an inflammatory amino acid shunt, and subsequent decreased expression of FH. FH inhibition further exacerbated fumarate accumulation and mitochondrial stress, while pharmacological inhibition of the shunt prevented this effect.
RNA sequencing and proteomics revealed that FH inhibition orchestrated a complex inflammatory response, suppressing the anti-inflammatory cytokine interleukin-10 (IL-10) and enhancing the pro-inflammatory cytokine tumor necrosis factor (TNF). This shift in cytokine balance was recapitulated by fumarate esters, suggesting that fumarate itself played a crucial role. The study also identified a novel mechanism by which FH regulated interferon production: by increasing the release and activation of mitochondrial RNA (mtRNA), which in turn activated immune sensors TLR7, RIG-I, and MDA5. This mtRNA-driven retrograde signaling pathway contributed to increased interferon-β (IFNβ) production, a key component of the innate immune response to viral infections.
The researchers further demonstrated that the effects of FH inhibition were not limited to the laboratory setting. In mice and humans, FH suppression was observed in response to prolonged LPS stimulation, indicating a potential role in regulating immune responses in vivo. Additionally, FH suppression was detected in patients with systemic lupus erythematosus (SLE), a chronic autoimmune disease characterized by excessive inflammation. This suggested that the FH-mtRNA axis might contribute to the pathogenesis of SLE and other inflammatory diseases.
Both studies identified mitochondrial dysfunction as a key event leading to innate immune activation. FH deficiency or inhibition led to impaired mitochondrial respiration, increased mitochondrial membrane potential, and mtDNA and mtRNA release into the cytosol. The findings from these studies have significant implications for understanding the pathogenesis of HLRCC and SLE and for developing new therapeutic strategies. The discovery that FH deficiency led to chronic inflammation and mtDNA release suggested a potential link between mtDNA release, chronic inflammation, and tumorigenesis in HLRCC. Targeting the pathways involved in mtDNA release or innate immune activation may offer new therapeutic opportunities for HLRCC treatment.
The identification of FH suppression in SLE patients and the role of mtRNA in IFNβ production raise the possibility that FH may be a novel therapeutic target for SLE. Restoring FH activity or inhibiting mtRNA release could potentially dampen the excessive IFNβ production observed in SLE, leading to improved disease outcomes.
Further research is needed to fully understand the mechanisms of mtDNA and mtRNA release and their role in innate immune activation. Additionally, investigating the interactions between metabolism, innate immunity, and other immune cells, such as T cells and B cells, is crucial for developing comprehensive models of immune responses in diseases like HLRCC and SLE.
Reference:
Zecchini V, Paupe V, Herranz-Montoya I, Janssen J, Wortel IMN, Morris JL, Ferguson A, Chowdury SR, Segarra-Mondejar M, Costa ASH, Pereira GC, Tronci L, Young T, Nikitopoulou E, Yang M, Bihary D, Caicci F, Nagashima S, Speed A, Bokea K, Baig Z, Samarajiwa S, Tran M, Mitchell T, Johnson M, Prudent J, Frezza C. Fumarate induces vesicular release of mtDNA to drive innate immunity. Nature. 2023 Mar;615(7952):499-506.
Hooftman A, Peace CG, Ryan DG, Day EA, Yang M, McGettrick AF, Yin M, Montano EN, Huo L, Toller-Kawahisa JE, Zecchini V, Ryan TAJ, Bolado-Carrancio A, Casey AM, Prag HA, Costa ASH, De Los Santos G, Ishimori M, Wallace DJ, Venuturupalli S, Nikitopoulou E, Frizzell N, Johansson C, Von Kriegsheim A, Murphy MP, Jefferies C, Frezza C, O'Neill LAJ. Macrophage fumarate hydratase restrains mtRNA-mediated interferon production. Nature. 2023 Mar;615(7952):490-498.






Post comments