
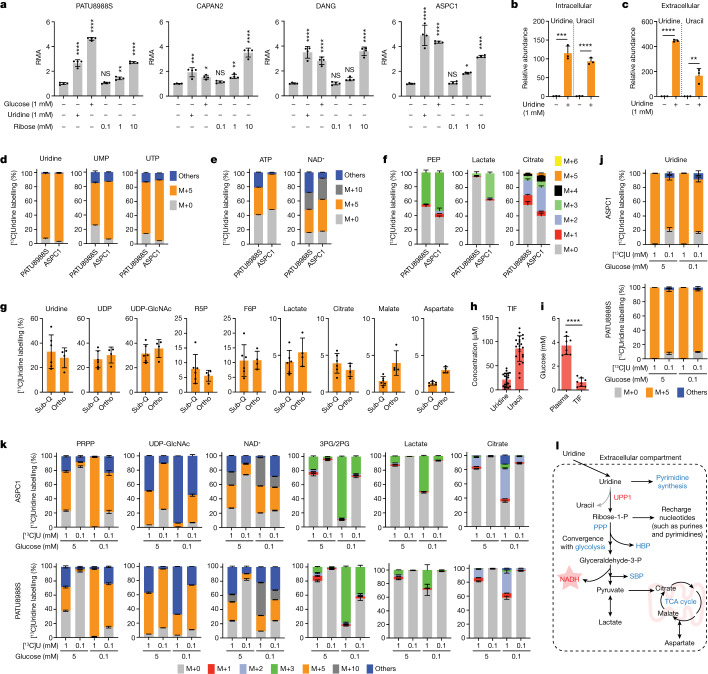
Credit:DOI:10.1038/s41586-023-06073-w. Profiling of metabolite utilization in PDA cells identifies uridine. a, Scheme of the nutrient metabolism screening assay and the correlation with gene expression in PDA cell lines and tumours. b, Spearman correlation (r) between the normalized relative metabolic activity (RMA) for uridine catabolism in the screening data and UPP1 mRNA expression data from an independent dataset (16 PDA cell lines). UPP1-high cell lines are shown in bold. c, The RMA in a subset of PDA cell lines following supplementation with 1 mM uridine for 3 days in glucose-free condition. d, Quantitative PCR (qPCR) validation of UPP1 mRNA expression in a subset of PDA cell lines. e, Immunoblot showing basal UPP1 expression in PDA cell lines. Blots are representative of three technical replicates with similar results. f, Spearman correlation (r) between protein densitometry analysis of the blot in e and UPP1 mRNA expression in the eight PDA cell lines highlighted in e. g, Top 20 genes differentially expressed by the PDA cell lines that were identified as uridine-high consumers/metabolizers compared with uridine-low consumers from the nutrient metabolism screen. Data source: Cancer Cell Line Encyclopedia (CCLE). Data in c,d are mean ± s.d.
Uridine is a nucleoside with an important role in metabolism. It serves as a source of energy, building blocks for nucleotides, and glucose. Uridine can be derived from dietary sources such as milk and beer, and it can be recycled from RNA. The uridine molecule is catabolized into ribose and uracil through the actions of uridine phosphorylase (UPP). Nwosu et al. identified uridine as a critical nutrient for the metabolism of pancreatic ductal adenocarcinoma (PDA) cells under nutrient-limiting conditions. The authors utilized a high-throughput Biolog phenotypic screening platform to screen over 175 metabolites for their impact on metabolic activity in 21 human PDA cell lines under nutrient restriction. The results revealed uridine as a fuel for PDA cells, and uridine utilization was found to correlate with the expression of UPP1. It was demonstrated that UPP1 liberated uridine-derived ribose to fuel central carbon metabolism, thereby supporting redox balance, survival, and proliferation in glucose-restricted PDA cells.
Consistent with the results in vitro, UPP1 expression was found to be elevated in PDA tumors compared to non-tumor tissues in mice, and high UPP1 expression correlated with poor survival in PDA patients. Furthermore, UPP1 deletion restricted the ability of PDA cells to use uridine and blunted tumor growth in mouse models. These findings identified uridine utilization as an important compensatory metabolic process in nutrient-deprived PDA cells, suggesting a novel metabolic axis for PDA therapy.
Notably, the expression and activity of UPP1 were found to be regulated by a complex interplay of cellular and molecular mechanisms. Mutations in the KRAS gene, which are common in pancreatic cancer, activate the KRAS-MAPK signaling pathway. This led to increased UPP1 expression, as evidenced by the higher levels of UPP1 observed in PDA tumors with the KRASG12D mutation compared to those without KRAS alterations. In addition, the pharmacological inhibition of MAPK pathway could suppress UPP1 expression and its activity.
In summary, this study provided a comprehensive understanding of the role of uridine in PDA metabolism and identified UPP1 as a key regulator of uridine utilization in PDA cells. The findings suggested that targeting the uridine-UPP1 axis may provide a novel therapeutic approach for the treatment of PDA. Further research is needed to validate the clinical significance of this metabolic vulnerability and to develop effective inhibitors of UPP1.
Another article, published in Nature Metabolism, also explored the uridine metabolism in 482 cancer cell lines and offer more information about its role in supporting energy production and cell growth in nutrient-limited conditions. The research conducted by Skinner et al. indicated that uridine could be metabolized into ribose, which then entered the pentose phosphate pathway (PPP) and glycolysis to provide energy and substrates for biosynthesis.
The study used a genome-wide genetic screen to identify genes required for uridine utilization in nutrient-limited conditions and uridine phosphorylase (UPP1/UPP2) emerged as a key gene. They demonstrated that cells expressing UPP1/UPP2 could grow in the absence of glucose, utilizing uridine for energy production. Metabolic tracer experiments confirmed the incorporation of uridine-derived carbon into glycolytic intermediates, indicating its utilization for energy production.
The study also demonstrated that uridine could support growth in not only cancer cell lines, but also primary macrophages and mice in vivo in the absence of glucose, indicating that the uridine-ribose metabolism pathway is widespread. Besides KRAS-MAPK signaling pathway mentioned in the first article, UPP1/UPP2 expression was found to be regulated by transcription factors such as MITF and NF-κB in the current study, suggesting that the pathway could be upregulated in response to diverse cellular signaling.
Furthermore, the authors suggested that this uridine-ribose metabolism pathway could be important in contexts such as mitochondrial dysfunction, immune response, and cancer metabolism. They also proposed that this pathway might be relevant to human diseases, as uridine is a common dietary constituent. The findings highlighted the role of uridine in energy production and cellular metabolism under nutrient-limited conditions, shedding light on potential therapeutic applications.
In addition to uridine, other nuleosides such as inosine, thymidine, and deoxycytidine have been identified as important metabolites in cancer. The upregulation of nucleoside usage under nutrient deprivation supports the idea that metabolic competition contributes to immunosuppression and tumor progression. These studies highlighted the significance of nucleosides in cancer metabolism.
Nwosu ZC, Ward MH, Sajjakulnukit P, Poudel P, Ragulan C, Kasperek S, Radyk M, Sutton D, Menjivar RE, Andren A, Apiz-Saab JJ, Tolstyka Z, Brown K, Lee HJ, Dzierozynski LN, He X, Ps H, Ugras J, Nyamundanda G, Zhang L, Halbrook CJ, Carpenter ES, Shi J, Shriver LP, Patti GJ, Muir A, Pasca di Magliano M, Sadanandam A, Lyssiotis CA. Uridine-derived ribose fuels glucose-restricted pancreatic cancer. Nature. 2023 Jun;618(7963):151-158.
Skinner OS, Blanco-Fernández J, Goodman RP, Kawakami A, Shen H, Kemény LV, Joesch-Cohen L, Rees MG, Roth JA, Fisher DE, Mootha VK, Jourdain AA. Salvage of ribose from uridine or RNA supports glycolysis in nutrient-limited conditions. Nat Metab. 2023 May;5(5):765-776.





Post comments